Quantitatively, plants are the only relevant primary producers on earth. Thus, they constitute the base for all life on the planet. However, production is restricted to the light phase, because light provides energy for reduction of CO2. During the night, plants themselfes consume reduced carbon to fuel maintenance of their organs.
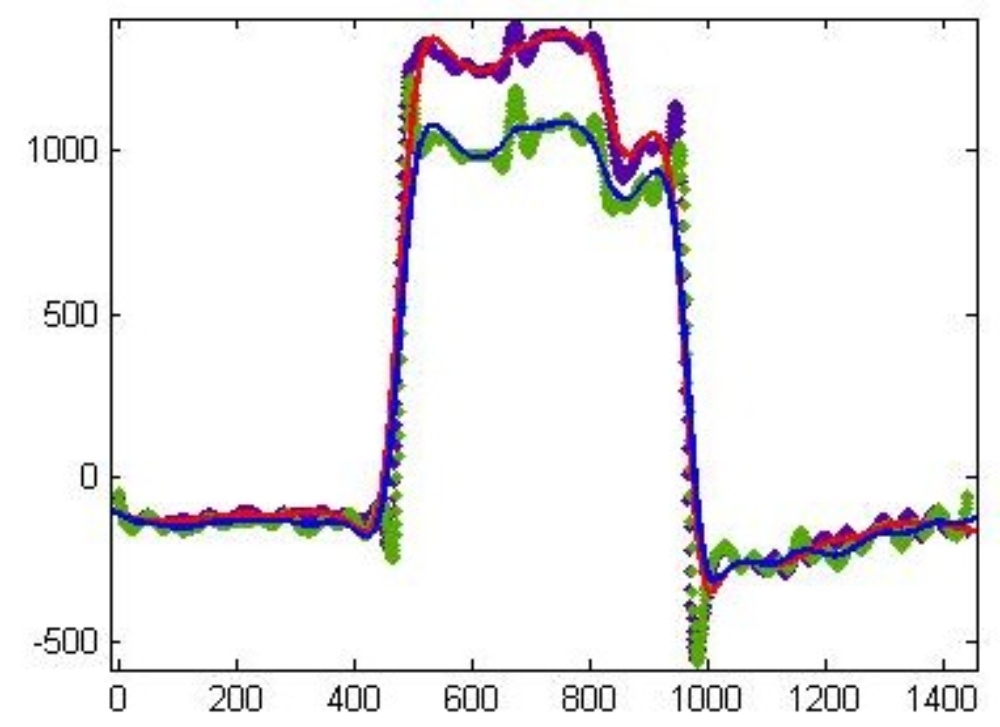
The figure on the right shows a diurnal profile of CO2 exchange of aerial organs in nmol/h*g fresh weight of an Arabidopsis thaliana plant during vegetative growth. The light phase starts at 8:00 am (480 min) and lasts until 4:00 pm (960 min). It becomse clear that during the light phase, more carbon is aquired than what is lost during the night. Thus, the aerial organs can export reduced carbon to supply sink organs like, e.g., the root. It is not easy to measure root respiration, because in soil, roots are associated with micro organisms that make a strong contribution to respiratory activity. We have developed a special technique to measure root respiration in an aeroponic system, and this allows us to study the full carbon balance of a plant over diurnal cycles.
During the light phase, plants produce carbohydrates that serve as building blocks for biomass and provide energy. Green leaves produce more carbohydrates than they need for their own metabolism, and they export the surplus to heterotrophic organs: young, growing leaves, roots that provide water and minerals, and flowers that asure reproduction.
Organs that export carbohydrates are called "sources", while those that import are called "sinks". Sources and sinks closely cooperate: only what is available can be consumed for growth and maintenance; but how much is produced also depends on demand.
We investigate, how supply and demand are regulated: do the sources dictate what will be available, or do they have to produce, what is needed?
In the relationship between sinks and sources, transport processes in the phloem, but also storage compounds play important roles. We investigate, if efficient storage of sugars in plant cells could stimulate photosynthesis. With respect to sugar storage, subcellular compartmentation is very important: while the large vacuole constitutes a nearly exhaustless reservoir, e.g., for fructans, storage of starch in the plastids can restrain photosynthesis. Nevertheless, in most plants, starch is the dominating carbon store. We investigate, how plants the detrmine the optimal amount of starch they produce to optimize diurnal change of light and dark phases.
Because carbohydrates function as transport as well as storage compounds, their diurnal dynamics are highly complex. To study organization of carbohydrate metabolism, we therefore use mathematical methods that allow simulation of non-intuitive processes.
A further level of complexity is given by the fact that carbohydrates also also involved in osmo-regulation of the cell. Thus, their concentrations are strongly influenced by environmental factors such as low temperature: during cold acclimation, large amounts of soluble sugars are accumulated that have protective functions. These environment interactions build another focus of our research.
Relevant Publications
- Friedrichs, N., Shokouhi, D., Heyer, A.G.: Flux Calculation for Primary Metabolism Reveals Changes in Allocation of Nitrogen to Different Amino Acid Families When Photorespiratory Activity Changes. International Journal of Molecular Sciences. 25, (2024). https://doi.org/10.3390/ijms25158394.
- Kraemer, K., Kepp, G., Brock, J., Stutz, S., Heyer, A.G.: Acclimation to elevated CO2 affects the C/N balance by reducing de novo N-assimilation. Physiologia Plantarum. 174, e13615 (2022). https://doi.org/10.1111/ppl.13615.
- Kraemer, K., Brock, J., Heyer, A.G.: Interaction of Nitrate Assimilation and Photorespiration at Elevated CO2. Frontiers in Plant Science. 13, (2022). https://doi.org/10.3389/fpls.2022.897924.
- Küstner, L., Nägele, T., Heyer, A.G.: Mathematical modeling of diurnal patterns of carbon allocation to shoot and root in Arabidopsis thaliana. npj Systems Biology and Applications. 5, 4–– (2019). https://doi.org/10.1038/s41540-018-0080-1.
- Küstner, L., Fürtauer, L., Weckwerth, W., Nägele, T., Heyer, A.G.: Subcellular dynamics of proteins and metabolites under abiotic stress reveal deferred response of the Arabidopsis thaliana hexokinase‐1 mutant gin2‐1 to high light. Plant Journal. 100, 456–472 (2019). https://doi.org/10.1111/tpj.14491.
- Fürtauer, L., Küstner, L., Weckwerth, W., Heyer, A.G., Nägele, T.: Resolving subcellular plant metabolism. Plant Journal. 100, 438–455 (2019). https://doi.org/10.1111/tpj.14472.
- Brauner, K., Birami, B., Brauner, H.A., Heyer, A.G.: Diurnal periodicity of assimilate transport shapes resource allocation and whole-plant carbon balance. The Plant Journal. 94, 776–789 (2018). https://doi.org/10.1111/tpj.13898.
- Birami, B., Gattmann, M., Heyer, A.G., Grote, R., Arneth, A., Ruehr, N.: Heat waves alter carbon allocation and increase mortality of Aleppo pine under dry conditions. Frontiers in Forests and Global Change. 1, 8 (2018). https://doi.org/doi: 10.3389/ffgc.2018.00008.
- Brauner, K., Stutz, S., Paul, M., Heyer, A.G.: Measuring whole plant CO2 exchange with the environment reveals opposing effects of the gin2–1 mutation in shoots and roots of <i>Arabidopsis thaliana</i>. Plant Signaling Behav. 10, e973822 (2015). https://doi.org/10.4161/15592324.2014.973822.
- Brauner, K., Hörmiller, I., Nägele, T., Heyer, A.G.: Exaggerated root respiration accounts for growth retardation in a starchless mutant of Arabidopsis thaliana. Plant J. 79, 82–91 (2014). https://doi.org/10.1111/tpj.12555.
- Nägele, T., Stutz, S., Hörmiller, I.I., Heyer, A.G.: Identification of a metabolic bottleneck for cold acclimation in Arabidopsis thaliana. Plant J. 72, 102–114 (2012). https://doi.org/10.1111/j.1365-313X.2012.05064.x.
- Zuther, E., Hoermiller, I.I., Heyer, A.G.: Evidence against sink limitation by the sucrose-to-starch route in potato plants expressing fructosyltransferases. Physiol. Plant. 143, 115–125 (2011). https://doi.org/10.1111/j.1399-3054.2011.01495.x.
- Wingenter, K., Trentmann, O., Winschuh, I., Hörmiller, I.I., Heyer, A.G., Reinders, J., Schulz, A., Geiger, D., Hedrich, R., Neuhaus, H.E.: A member of the mitogen-activated protein 3-kinase family is involved in the regulation of plant vacuolar glucose uptake. Plant J. 68, 890–900 (2011). https://doi.org/10.1111/j.1365-313X.2011.04739.x.
- Henkel, S., Nägele, T., Hörmiller, I., Sauter, T., Sawodny, O., Ederer, M., Heyer, A.G.: A systems biology approach to analyse leaf carbohydrate metabolism in Arabidopsis thaliana. EURASIP Journal on Bioinformatics and Systems Biology. 2011, 2 (2011). https://doi.org/10.1186/1687-4153-2011-2.
- Wingenter, K., Schulz, A., Wormit, A., Wic, S., Trentmann, O., Hoermiller, I.I., Heyer, A.G., Marten, I., Hedrich, R., Neuhaus, H.E.: Increased Activity of the Vacuolar Monosaccharide Transporter TMT1 Alters Cellular Sugar Partitioning, Sugar Signaling, and Seed Yield in Arabidopsis. Plant Physiol. 154, 665–677 (2010). https://doi.org/10.1104/pp.110.162040.
- Nägele, T., Henkel, S., Hörmiller, I., Sauter, T., Sawodny, O., Ederer, M., Heyer, A.G.: Mathematical modeling of the central carbohydrate metabolism in Arabidopsis reveals a substantial regulatory influence of vacuolar invertase on whole plant carbon metabolism. Plant Physiol. 153, 260–272 (2010). https://doi.org/10.1104/pp.110.154443.
- Livingston, D.P., Hincha, D.K., Heyer, A.G.: Fructan and its relationship to abiotic stress tolerance in plants. Cellular and Molecular Life Sciences. 66, 2007–2023 (2009). https://doi.org/10.1007/s00018-009-0002-x.
- Hincha, D.K., Livingston, D.P., Premakumar, R., Zuther, E., Obel, N., Cacela, C., Heyer, A.G.: Fructans from oat and rye: Composition and effects on membrane stability during drying. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1768, 1611–1619 (2007). https://doi.org/10.1016/j.bbamem.2007.03.011.
- Hannah, M.A., Kramer, K.M., Geffroy, V., Kopka, J., Blair, M.W., Erban, A., Vallejos, C.E., Heyer, A.G., Sanders, F.E.T., Millner, P.A., Pilbeam, D.J.: Hybrid weakness controlled by the dosage-dependent lethal (DL) gene system in common bean (Phaseolus vulgaris) is caused by a shoot-derived inhibitory signal leading to salicylic acid-associated root death. New Phytol. 176, 537–549 (2007). https://doi.org/10.1111/j.1469-8137.2007.02215.x.
- Hannah, M.A., Zuther, E., Buchel, K., Heyer, A.G.: Transport and metabolism of raffinose family oligosaccharides in transgenic potato. J. Exp. Bot. 57, 3801–3811 (2006). https://doi.org/10.1093/jxb/erl152.
- Zuther, E., Kwart, M., Willmitzer, L., Heyer, A.G.: Expression of a yeast-derived invertase in companion cells results in long-distance transport of a trisaccharide in an apoplastic loader and influences sucrose transport. Planta. 218, 759–766 (2004). https://doi.org/10.1007/s00425-003-1148-7.
- Heyer, A.G., Raap, M., Schroeer, B., Marty, B., Willmitzer, L.: Cell wall invertase expression at the apical meristem alters floral, architectural, and reproductive traits in Arabidopsis thaliana. Plant Journal. 39, 161–169 (2004). https://doi.org/10.1111/j.1365-313X.2004.02124.x.
- Heyer, A.G., Wendenburg, R.: Gene Cloning and Functional Characterization by Heterologous Expression of the Fructosyltransferase of Aspergillus sydowi IAM 2544. Applied and Environmental Microbiology. 67, 363–370 (2001). https://doi.org/10.1128/AEM.67.1.363-370.2001.
- Wolff, D., Czapla, S., Heyer, A.G., Radosta, S., Mischnick, P., Springer, J.: Globular shape of high molar mass inulin revealed by static light scattering and viscometry. Polymer. 41, 8009–8016 (2000). https://doi.org/10.1016/s0032-3861(00)00168-3.
- Hellwege, E.M., Czapla, S., Jahnke, A., Willmitzer, L., Heyer, A.G.: Transgenic potato (Solanum tuberosum) tubers synthesize the full spectrum of inulin molecules naturally occurring in globe artichoke (Cynara scolymus) roots. Proceedings of the National Academy of Sciences. 97, 8699–8704 (2000). https://doi.org/10.1073/pnas.150043797.